Кандидат биологических наук Л. Чайлахян, научный сотрудник Института биофизики АН СССР
Читательница журнала Л. Горбунова (деревня Цыбино, Московской области) пишет нам: «Меня интересует механизм, передачи сигналов по нервным, клеткам».
Лауреаты Нобелевской премии 1963 года (слева направо): А. Ходжкин, Э. Хаксли, Д. Экклс.
Представления ученых о механизме передачи нервного импульса претерпели в последнее время существенное изменение. До недавнего времени в науке господствовали взгляды Бернштейна.
Мозг человека, без сомнения, высшее достижение природы. В килограмме нервной ткани заключена квинтэссенция всего человека, начиная от регуляции жизненных функций - работы сердца, легких, пищеварительного тракта, печени - и кончая его духовным миром. Здесь - наши мыслительные способности, всё наше мироощущение, память, разум, наше самосознание, наше «я». Познание механизмов работы мозга - это познание самого себя.
Велика и заманчива цель, но неимоверно сложен объект исследования. Шутка сказать, этот килограмм ткани представляет собой сложнейшую систему связи десятков миллиардов нервных клеток.
Однако первый существенный шаг к познанию работы мозга уже сделан. Может быть, он один из самых легких, но он чрезвычайно важен для всего дальнейшего.
Я имею в виду исследование механизма передачи нервных импульсов - сигналов, бегущих по нервам, как по проводам. Именно эти сигналы являются той азбукой мозга, с помощью которой органы чувств посылают в центральную нервную систему сведения-депеши о событиях во внешнем мире. Нервными импульсами зашифровывает мозг свои приказы мышцам и различным внутренним органам. Наконец, на языке этих сигналов говорят между собой отдельные нервные клетки и нервные центры.
Нервные клетки - основной элемент мозга - разнообразны по величине, по форме, но в принципе обладают единым строением. Каждая нервная клетка состоит из трех частей: из тела, длинного нервного волокна - аксона (длина его у человека от нескольких миллиметров до метра) и нескольких коротких ветвистых отростков - дендритов. Нервные клетки изолированы друг от друга оболочками. Но все же клетки взаимодействуют между собой. Происходит это в месте стыка клеток; этот стык называется «синапс». В синапсе встречаются аксон одной нервной клетки и тело или дендрит другой клетки. Причем интересно, что возбуждение может передаваться только и одном направлении: от аксона к телу или дендриту, но ни в коем случае не обратно. Синапс - это как бы кенотрон: он пропускает сигналы только в одном направлении.
В проблеме изучения механизма нервного импульса и его распространения можно выделить два основных вопроса: природа проведения нервного импульса или возбуждения в пределах одной клетки - по волокну и механизм передачи нервного импульса от клетки к клетке - через синапсы.
Какова природа сигналов, передающихся от клетки к клетка по нервным волокнам?
Этой проблемой человек интересовался уже давно, Декарт предполагал, что распространение сигнала связано с переливанием жидкости по нервам, как по трубкам. Ньютон думал, что это чисто механический процесс. Когда появилась электромагнитная теория, ученые решили, что нервный импульс аналогичен движению тока по проводнику со скоростью, близкой к скорости распространения электромагнитных колебаний. Наконец, с развитием биохимии появилась точка зрения, что движение нервного импульса - это распространение вдоль по нервному волокну особой биохимической реакции.
И всё же ни одно из этих представлений не оправдалось.
В настоящее время природа нервного импульса раскрыта: это удивительно тонкий электрохимический процесс, в основе которого лежит перемещение ионов через оболочку клетки.
Большой вклад в раскрытие этой природы внесли работы трех ученых: Алана Ходжкина, профессора биофизики Кембриджского университета; Эндрью Хаксли, профессора физиологии Лондонского университета, и Джона Экклса, профессора физиологии австралийского университета в Канберре. Им присуждена Нобелевская премия в области медицины за 1963 год,
Впервые предположение об электрохимической природе нервного импульса высказал известный немецкий физиолог Бернштейн в начале нашего столетия.
К началу двадцатого века было довольно многое известно о нервном возбуждении. Ученые уже знали, что нервное волокно можно возбудить электрическим током, причем возбуждение всегда возникает под катодом - под минусом. Было известно, что возбужденная область нерва заряжается отрицательно по отношению к невозбужденному участку. Было установлено, что нервный импульс в каждой точке длится всего 0,001-0,002 секунды, что величина возбуждения не зависит от силы раздражения, как громкость звонка в нашей квартире не зависит от того, как сильно мы нажимаем на кнопку. Наконец, ученые установили, что носителями электрического тока в живых тканях являются ионы; причем внутри клетки основной электролит - соли калия, а в тканевой жидкости - соли натрия. Внутри большинства клеток концентрация ионов калия в 30-50 раз больше, чем в крови и в межклеточной жидкости, омывающей клетки.
И вот на основании всех этих данных Бернштейн предположил, что оболочка нервных и мышечных клеток представляет собой особую полупроницаемую мембрану. Она проницаема только для ионов К + ; для всех остальных ионов, в том числе и для находящихся внутри клетки отрицательно заряженных анионов, путь закрыт. Ясно, что калий по законам диффузии будет стремиться выйти из клетки, в клетке возникает избыток анионов, и по обе стороны мембраны появится разность потенциалов: снаружи - плюс (избыток катионов), внутри - минус (избыток анионов). Эта разность потенциалов получила название потенциала покоя. Таким образом, в покое, в невозбужденном состоянии внутренняя часть клетки всегда заряжена отрицательно по сравнению с наружным раствором.
Бернштейн предположил, что в момент возбуждения нервного волокна происходят структурные изменения поверхностной мембраны, ее поры как бы увеличиваются, и она становится проницаемой для всех ионов. При этом, естественно, разность потенциалов исчезает. Это и вызывает нервный сигнал.
Мембранная теория Бернштейма быстро завоевала признание и просуществовала свыше 40 лет, вплоть до середины нашего столетия.
Но уже в конце 30-х годов теория Бернштейна встретилась с непреодолимыми противоречиями. Сильный удар ей был нанесен в 1939 году тонкими экспериментами Ходжкина и Хаксли. Эти ученые впервые измерили абсолютные величины мембранного потенциала нервного волокна в покое и при возбуждении. Оказалось, что при возбуждении мембранный потенциал не просто уменьшался до нуля, а переходил через ноль на несколько десятков милливольт. То есть внутренняя часть волокна из отрицательной становилась положительной.
Но мало ниспровергнуть теорию, надо заменить ее другой: наука не терпит вакуума. И Ходжкин, Хаксли, Катц в 1949-1953 годах предлагают новую теорию. Она получает название натриевой.
Здесь читатель вправе удивиться: до сих пор о натрии не было речи. В этом все и дело. Ученые установили с помощью меченых атомов, что в передаче нервного импульса замешаны не только ионы калия и анионы, но и ионы натрия и хлора.
В организме достаточно ионов натрия и хлора, все знают, что кровь соленая на вкус. Причем натрия в межклеточной жидкости в 5-10 раз больше, чем внутри нервного волокна.
Что же это может означать? Ученые предположили, что при возбуждении в первый момент резко увеличивается проницаемость мембраны только для натрия. Проницаемость становится в десятки раз больше, чем для ионов калия. А так как натрия снаружи в 5-10 рез больше, чем внутри, то он будет стремиться войти в нервное волокно. И тогда внутренняя часть волокна станет положительной.
А через какое-то время - после возбуждения - равновесие восстанавливается: мембрана начинает пропускать и ионы калия. И они выходят наружу. Тем самым они компенсируют тот положительный заряд, который был внесен внутрь волокна ионами натрия.
Совсем нелегко было прийти к таким представлениям. И вот почему: диаметр иона натрия в растворе раза в полтора больше диаметра ионов калия и хлора. И совершенно непонятно, каким образом больший по размеру ион проходит там, где не может пройти меньший.
Нужно было решительно изменить взгляд на механизм перехода ионов через мембраны. Ясно, что только рассуждениями о порах в мембране здесь не обойтись. И тогда была высказана идея, что ионы могут пересекать мембрану совершенно другим способом, с помощью тайных до поры до времени союзников - особых органических молекул-переносчиков, спрятанных в самой мембране. С помощью такой молекулы ионы могут пересекать мембрану в любом месте, а не только через поры. Причем эти молекулы-такси хорошо различают своих пассажиров, они не путают ионы натрия с ионами калия.
Тогда общая картина распространения нервного импульса будет иметь следующий вид. В покое молекулы-переносчики, заряженные отрицательно, мембранным потенциалом прижаты к наружной границе мембраны. Поэтому проницаемость для натрия очень мала: в 10-20 раз меньше, чем для ионов калия. Калий может пересекать мембрану через поры. При приближении волны возбуждения уменьшается давление электрического поля на молекулы-переносчики; они сбрасывают свои электростатические «оковы» и начинают переносить ионы натрия внутрь клетки. Это еще больше уменьшает мембранный потенциал. Идет как бы цепной процесс перезарядки мембраны. И этот процесс непрерывно распространяется вдоль нервного волокна.
Интересно, что нервные волокна тратят на свою основную работу - проведение нервных импульсов - всего около 15 минут в сутки. Однако готовы к этому волокна в любую секунду: все элементы нервного волокна работают без перерыва - 24 часа в сутки. Нервные волокна в этом смысле подобны самолетам-перехватчикам, у которых непрерывно работают моторы для мгновенного вылета, однако сам вылет может состояться лишь раз в несколько месяцев.
Мы познакомились сейчас с первой половиной таинственного акта прохождения нервного импульса - вдоль одного волокна. А как же передается возбуждение от клетки к клетке, через места стыков - синапсы. Этот вопрос был исследован в блестящих опытах третьего нобелевского лауреата, Джона Экклса.
Возбуждение не может непосредственно перейти с нервных окончаний одной клетки на тело или дендриты другой клетки. Практически весь ток вытекает через синаптическую щель в наружную жидкость, и в соседнюю клетку через синапс попадает ничтожная его доля, неспособная вызвать возбуждение. Таким образом, в области синапсов электрическая непрерывность в распространении нервного импульса нарушается. Здесь, на стыке двух клеток, в силу вступает совершенно другой механизм.
Когда возбуждение подходит к окончанию клетки, к месту синапса, в межклеточную жидкость выделяются физиологически активные вещества - медиаторы, или посредники. Они становятся связующим звеном в передаче информации от клетки к клетке. Медиатор химически взаимодействует со второй нервной клеткой, изменяет ионную проницаемость ее мембраны - как бы пробивает брешь, в которую устремляются многие ионы, в том числе и ионы натрия.
Итак, благодаря работам Ходжкина, Хаксли и Экклса важнейшие состояния нервной клетки - возбуждение и торможение - можно описать в терминах ионных процессов, в терминах структурно-химических перестроек поверхностных мембран. На основании этих работ уже можно делать предположения о возможных механизмах кратковременной и долговременной памяти, о пластических свойствах нервной ткани. Однако это разговор о механизмах в пределах одной или нескольких клеток. Это лишь, азбука мозга. По-видимому, следующий этап, возможно, гораздо более трудный, - вскрытие законов, по которым строится координирующая деятельность тысяч нервных клеток, распознание языка, на котором говорят между собой нервные центры.
Мы сейчас в познании работы мозга находимся на уровне ребенка, который узнал буквы алфавита, но не умеет связывать их в слова. Однако недалеко время, когда ученые с помощью кода - элементарных биохимических актов, происходящих в нервной клетке, прочтут увлекательнейший диалог между нервными центрами мозга.
Детальное описание иллюстраций
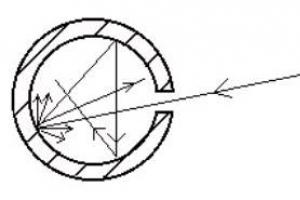
Представления ученых о механизме передачи нервного импульса претерпели в последнее время существенное изменение. До недавнего времени в науке господствовали взгляды Бернштейна. По его мнению, в состоянии покоя (1) нервное волокно заряжено положительно снаружи и отрицательно внутри. Это объяснялось тем, что сквозь поры в стенке волокна могут проходить только положительно заряженные ионы калия (К +); большие по размерам отрицательно наряженные анионы (А –) вынуждены оставаться внутри и создавать избыток отрицательных зарядов. Возбуждение (3) по Бернштейну сводится к исчезновению разности потенциалов, которое вызывается тем, что размер пор увеличивается, анионы выходят наружу и выравнивают ионный баланс: количество положительных ионов становится равным количеству отрицательных. Работа лауреатов Нобелевской премии 1963 года А. Ходжкпна, Э. Хаксли и Д. Экклса изменила наши прежние представления. Доказано, что в нервном возбуждении участвуют также положительные ионы натрия (Na +), отрицательные ноны хлора (Сl –) и отрицательно заряженные молекулы-переносчики. Покоящееся состояние (3) образуется в принципе так же, как и считалось раньше: избыток положительных ионов - снаружи нервного волокна, избыток отрицательных - внутри. Однако установлено, что при возбуждении (4) происходит не выравнивание зарядов, а перезарядка: снаружи образуется избыток отрицательных ионов, а внутри - избыток положительных. Объясняется это тем, что при возбуждении молекулы-переносчики начинают перевозить сквозь стенку положительные ионы натрия. Таким образом, нервный импульс (5) - это перемещающаяся вдоль волокна перезарядка двойного электрического слоя. А от клетки к клетке возбуждение передается своеобразным химическим «тараном» (6) - молекулой ацетилхолина, которая помогает ионам прорываться сквозь стенку соседнего нервного волокна.
НЕРВНЫЙ ИМПУЛЬС (лат. nervus нерв; лат. impulsus удар, толчок) - волна возбуждения, распространяющаяся по нервному волокну; единица распространяющегося возбуждения.
Н. и. обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам - скелетной мускулатуре, гладким мышцам внутренних органов и сосудов, железам внутренней и внешней секреции и т. д.
Сложная информация о действующих на организм раздражениях кодируется в виде отдельных групп Н. и.- рядов. Согласно закону «Все или ничего » (см.) амплитуда и длительность отдельных Н. и., проходящих по одному и тому же волокну, постоянны, а частота и количество Н. и. в ряду зависят от интенсивности раздражения. Такой способ передачи информации является наиболее помехоустойчивым, т. е. в широких пределах не зависит от состояния проводящих волокон.
Распространение Н. и. отождествляется с проведением потенциалов действия (см. Биоэлектрические потенциалы). Возникновение возбуждения может быть результатом раздражения (см.), напр, воздействие света на зрительный рецептор, звука на слуховой рецептор, или процессов, протекающих в тканях (спонтанное возникновение Н. и.). В этих случаях Н. и. обеспечивают согласованную работу органов при протекании какого-либо физиол, процесса (напр., в процессе дыхания Н. и. вызывают сокращение скелетных мышц и диафрагмы, результатом чего являются вдох и выдох, и т. д.).
В живых организмах передача информации может осуществляться и гуморальным путем, посредством выброса в русло крови гормонов, медиаторов и т. п. Однако преимущество информации, передаваемой при помощи Н. и., состоит в том, что она более целенаправленна, передается быстро и может быть точнее закодирована, чем сигналы, посылаемые гуморальной системой.
Факт, что нервные стволы являются путем, по к-рому передаются влияния от мозга к мышцам и в обратном направлении, был известен еще в эпоху античности. В средние века и вплоть до середины 17 в. считалось, что по нервам распространяется некая субстанция, подобная жидкости или пламени. Идея о электрической природе Н. и. возникла в 18 в. Первые исследования электрических явлений в живых тканях, связанных с возникновением и распространением возбуждения, были осуществлены Л. Галь-вани. Г. Гельмгольц показал, что скорость распространения Н. и., к-рую ранее считали близкой к скорости света, имеет конечное значение и может быть точно измерена. Германн (L. Hermann) ввел в физиологию понятие потенциала действия. Объяснение механизма возникновения и проведения возбуждения стало возможным после создания С. Аррениусом теории электролитической диссоциации. В соответствии с этой теорией Бернштейн (J. Bernstein) предположил, что возникновение и проведение Н. и. обусловлено перемещением ионов между нервным волокном и окружающей средой. Англ. исследователи А. Ходжкин, Б. Катц и Э. Хаксли детально исследовали трансмембранные ионные токи, лежащие в основе развития потенциала действия. Позже стали интенсивно изучаться механизмы работы ионных каналов, по к-рым происходит обмен ионами между аксоном и окружающей средой, и механизмы, обеспечивающие способность нервных волокон проводить ряды Н. и. разного ритма и продолжительности.
Н. и. распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками нервного волокна. Ток, выходящий из волокна наружу в покоящемся участке, служит раздражителем. Наступающая после возбуждения в данном участке нервного волокна рефрактерность обусловливает поступательное движение Н. и.
Количественно соотношения разных фаз развития потенциала действия можно охарактеризовать, сопоставляя их по амплитуде и длительности во времени. Так, напр., для миелиновых нервных волокон группы А млекопитающих диаметр волокна находится в пределах 1-22 мк, скорость проведения - 5-120 м/сек, длительность и амплитуда высоковольтной части (пика, или спайка) - 0,4-0,5 мсек и 100-120 мв соответственно, следовой негативный потенциал - 12-20 мсек (3-5% от амплитуды спайка), следовой позитивный потенциал - 40-60 мсек (0,2% от амплитуды спайка).
Возможности передачи разнообразной информации расширяются за счет повышения скорости развития потенциала действия, скорости распространения, а также за счет повышения лабильности (см.) - т. е. способности возбудимого образования воспроизводить в единицу времени высокие ритмы возбуждения.
Конкретные особенности распространения Н. и. связаны со строением нервных волокон (см.). Сердцевина волокна (аксоплазма) обладает низким сопротивлением и, соответственно, хорошей проводимостью, а окружающая аксоплазму плазматическая мембрана - большим сопротивлением. Особенно велико электрическое сопротивление наружного слоя у миелинизированных волокон, у к-рых свободны от толстой миелиновой оболочки только перехваты Ранвье. В безмиелиновых волокнах Н. и. движется непрерывно, а в миелиновых - скачкообразно (сальтаторное проведение).
Различают декрементное и бездекрементное распространение волны возбуждения. Декрементное проведение, т. е. проведение возбуждения с угасанием, наблюдается в безмиелиновых волокнах. В таких волокнах скорость проведения Н. и. невелика и по мере отдаления от места раздражения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания. Декрементное проведение свойственно волокнам, иннервирующим внутренние органы, обладающие низкой функц, подвижностью. Без декрементное проведение характерно для миелиновых и тех безмиелиновых волокон, к-рые передают сигналы к органам, обладающим высокой реактивностью (напр., сердечной мышце). При бездекрементном проведении Н. и. проходит весь путь от места раздражения до места реализации информации без затухания.
Максимальная скорость проведения Н. и., зарегистрированная в быстропроводящих нервных волокнах млекопитающих, составляет 120 м/сек. Высокие скорости проведения импульса могут быть достигнуты за счет увеличения диаметра нервного волокна (у безмиелиновых волокон) или за счет повышения степени миелинизации. Распространение одиночного Н. и. само по себе не требует непосредственных энергетических затрат, т. к. при определенном уровне поляризации мембраны каждый участок нервного волокна находится в состоянии готовности к проведению и раздражающий стимул играет роль «спускового курка». Однако восстановление исходного состояния нервного волокна и поддержание его в готовности к проведению нового Н. и. связано с затратой энергии биохим, реакций, протекающих в нервном волокне. Процессы восстановления приобретают большое значение в случае проведения рядов Н. и. При проведении ритмического возбуждения (рядов импульсов) в нервных волокнах приблизительно вдвое возрастает теплопродукция и потребление кислорода, расходуются макроэргические фосфаты и повышается активность Na,K-АТФ-азы к-рую отождествляют с натриевым насосом. Изменение интенсивности протекания различных физ.-хим. и биохим, процессов зависит от характера ритмического возбуждения (продолжительность рядов импульсов и частота их следования) и физиол, состояния нерва. При проведении большого числа Н. и. в высоком ритме в нервных волокнах может накапливаться «метаболический долг» (это находит отражение в увеличении суммарных следовых потенциалов), и тогда процессы восстановления затягиваются. Но и в этих условиях способность нервных волокон проводить Н. и. долгое время остается неизменной.
Передача Н. и. с нервного волокна на мышечное или какой-либо другой эффектор осуществляется через синапсы (см.). У позвоночных животных в подавляющем большинстве случаев передача возбуждения на эффектор происходит при помощи выделения ацетилхолина (нервно-мышечные синапсы скелетной мускулатуры, синаптические соединения в сердце и др.). Для таких синапсов характерно строго одностороннее проведение импульса и наличие временной задержки передачи возбуждения.
В синапсах, в синаптической щели которых сопротивление электрическому току благодаря большой площади контактирующих поверхностей мало, происходит электрическая передача возбуждения. В них нет синаптической задержки проведения и возможно двустороннее проведение. Такие синапсы свойственны беспозвоночным животным.
Регистрация Н. и. нашла широкое применение в биол, исследованиях и клин, практике. Для регистрации используют шлейфные и чаще катодные осциллографы (см. Осциллография). При помощи микроэлектродной техники (см. Микроэлектродный метод исследования) регистрируют Н. и. в одиночных возбудимых образованиях - нейронах и аксонах. Возможности исследования механизма возникновения и распространения Н. и. значительно расширились после разработки метода фиксации потенциала. Этим методом были получены основные данные о ионных токах (см. Биоэлектрические потенциалы).
Нарушение проведения Н. и. происходит при повреждении нервных стволов, напр, при механических травмах, сдавливании в результате разрастания опухоли или при воспалительных процессах. Такие нарушения проведения Н. и. зачастую бывают необратимы. Следствием прекращения иннервации могут быть тяжелые функциональные и трофические расстройства (напр., атрофия скелетных мышц конечностей после прекращения поступления Н. и. вследствие необратимой травмы нервного ствола). Обратимое прекращение проведения Н. и. может быть вызвано специально, в терапевтических целях. Напр., с помощью анестезирующих средств блокируют импульсацию, идущую от болевых рецепторов в ц. н. с. Обратимое прекращение проведения Н. и. вызывает и новокаиновая блокада. Временное прекращение передачи Н. и. по нервным проводникам наблюдается и во время общего наркоза.
Библиография: Бpеже М. А. Электрическая активность нервной системы, пер. с англ., М., 1979; Жуков Е. К. Очерки по нервно-мышечной физиологии, JI., 1969; К о н н e л и К. Восстановительные процессы и обмен веществ в нерве, в кн.: Совр, пробл. биофизики, пер. с англ., под ред. Г. М. Франка и А. Г. Па-сынского, т. 2, с. 211, М., 1961;
Костюк П. Г. Физиология центральной нервной системы, Киев, 1977; Л а т м а-низова JI. В. Очерк физиологии возбуждения, М., 1972; Общая физиология нервной системы, под ред. П. Г. Костюка, JI., 1979; T а с а к и И. Нервное возбуждение, пер. с англ., М., 1971; Ходжкин А. Нервный импульс, пер. с англ., М., 1965; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975.
НЕРВНЫЙ ИМПУЛЬС
НЕРВНЫЙ ИМПУЛЬС
Волна возбуждения, к-рая распространяется по нервному волокну и служит для передачи информации от периферич. рецепторных (чувствительных) окончаний к нервным центрам, внутри центр. нервной системы и от неё к исполнительным аппаратам - мышцам и железам. Прохождение Н. и. сопровождается переходными электрич. процессами, к-рые можно зарегистрировать как внеклеточными, так и внутриклеточными электродами.
Генерацию, передачу и переработку Н. и. осуществляет нервная система. Осн. структурным элементом нервной системы высших организмов является нервная клетка, или нейрон, состоящий из тела клетки и многочисл. отростков - дендритов (рис. 1). Один из отростков у нериферич. нейронов имеет большую длину - это нервное волокно, или аксон, протяжённость к-рого ~ 1 м, а толщина от 0,5 до 30 мкм. Различают два класса нервных волокон: мякотные (мие-линизированные) и безмякотные. У мякотных волокон имеется миелиновая , образованная спец. мембраной, к-рая подобно изоляции накручивается на аксон. Протяжённость участков сплошной миелиновой оболочки составляет от 200 мкм до 1 мм, они прерываются т. н. перехватами Ранвье шириной 1мкм. Миелиновая оболочка играет роль изоляции; нервное волокно на этих участках пассивно, электрически активна только в перехватах Ранвье. Безмякотные волокна не имеют изолир. участков; их структура однородна по всей длине, а мембрана обладает электрич. активностью по всей поверхности.
Нервные волокна заканчиваются на телах или ден-дритах др. нервных клеток, но отделены от них проме-
жутком шириной ~ 10 нм. Эта область контакта двух клеток наз.
синапсом. Входящая в синапс мембрана аксона наз. пресинаптической, а соответствующая мембрана дендритов или мышцы - пост-синаптической (см. Клеточные структуры).
В нормальных условиях по нервному волокну постоянно бегут серии Н. и., возникающих на дендритах или теле клетки и распространяющихся по аксону в направлении от тела клетки (аксон может проводить Н. и. в обоих направлениях). Частота этих периодич. разрядов несёт информацию о силе вызвавшего их раздражения; напр., при умеренной активности частота ~ 50-100 импульсов/с. Существуют клетки, к-рые разряжаются с частотой ~ 1500 импульсов/с.
Скорость распространения Н. и. u.
зависит от типа нервного волокна и его диаметра d,
u.
~ d
1/2 . В тонких волокнах нервной системы человека u.
~ 1 м/с, а в толстых волокнах u.
~ 100-120 м/с.
Каждый Н. и. возникает в результате раздражения тела нервной клетки или нервного волокна. Н. и. всегда имеет одни и те же характеристики (форму и скорость) независимо от силы раздражения, т. е. при подпороговом раздражении Н. и. не возникает совсем, а при надпороговым - имеет полную амплитуду.
После возбуждения наступает рефракторный период, в течение к-рого возбудимость нервного волокна снижена. Различают абс. рефракторный период, когда волокно нельзя возбудить никакими раздражителями, и относит. рефракторный период, когда возможно, но его порог оказывается выше нормы. Абс. рефракторный период ограничивает сверху частоту передачи Н. и. Нервное волокно обладает свойством аккомодации, т. е. привыкает к постоянно действующему раздражению, что выражается в постепенном повышении порога возбудимости. Это приводит к снижению частоты Н. и. и даже к их полному исчезновению. Если раздражения нарастает медленно, то возбуждения может не произойти даже после достижения порога.
Рис.1. Схема строения нервной клетки.
Вдоль нервного волокна Н. и. распространяется в виде электрич. потенциала. В синапсе происходит смена механизма распространения. Когда Н. и. достигает пресинаптич. окончания, в синаптич. щель выделяется активное хим. - м е д и а т о р. Медиатор диффундирует через синаптич. щель и меняет проницаемость постсинаптич. мембраны, в результате чего на ней возникает , вновь генерирующий распространяющийся . Так действует хим. синапс. Встречается также электрич. синапс, когда . нейрон возбуждается электрически.
Возбуждение Н. и.
Физ. представления о появлении электрич. потенциалов в клетках основаны на т. н. мембранной теории. Клеточные мембраны разделяют электролита разной концентрации и обладают из-бират. проницаемостью для нек-рых ионов. Так, мембрана аксона представляет собой тонкий слой липидов и белков толщиной ~ 7 нм. Её электрич. сопротивление в состоянии покоя ~ 0,1 Ом. м 2 , а ёмкость ~ 10 мф/м 2 . Внутри аксона высока ионов К + и мала концентрация ионов Na + и Сl - , а в окружающей среде - наоборот.
В состоянии покоя мембрана аксона проницаема для ионов К + . Из-за разницы концентраций C 0 K .
во внеш. и С во внутр. растворах на мембране устанавливается калиевый мембранный потенциал
![]()
где Т -
абс. темп-pa, е -
заряд электрона. На мембране аксона действительно наблюдается потенциал покоя ~ -60 мВ, соответствующий указанной ф-ле.
Ионы Na + и Сl - проникают через мембрану. Для поддержания необходимого неравновесного распределения ионов клетка использует систему активного транспорта, на работу к-рой расходуется клеточная . Поэтому состояние покоя нервного волокна не является термодинамически равновесным. Оно стационарно благодаря действию ионных насосов, причём мембранный потенциал в условиях разомкнутой цепи определяется из равенства нулю полного электрич. тока.
Процесс нервного возбуждения развивается следующим образом (см. также Биофизика).
Если пропустить через аксон слабый импульс тока, приводящий к деполяризации мембраны, то после снятия внеш. воздействия потенциал монотонно возвращается к исходному уровню. В этих условиях аксон ведёт себя как пассивная электрич. цепь, состоящая из конденсатора и пост. сопротивления.
Рис. 2. Развитие потенциала действия в нервном во
локне: а
- подпороговое (1
) и надпороговое (2)
раздражения; б
-мембранный отклик; при над-пороговом раздражении проявляется полный потен
циал действия; в
- ионный ток, протекающий через
мембрану при возбуждении; г -
аппроксимация
ионного тока в простой аналитической модели.

Если импульс тока превышает нек-рую пороговую величину, потенциал продолжает изменяться и после выключения возмущения; потенциал становится положительным и только потом возвращается к уровню покоя, причём вначале даже несколько проскакивает его (область гиперполяризации, рис. 2). Отклик мембраны при этом не зависит от возмущения; этот импульс наз. потенциалом действия. Одновременно через мембрану течёт ионный ток, направленный сначала внутрь, а потом наружу (рис. 2, в
).
Феноменологич. истолкование механизма возникновения Н. и. было дано А. Л. Ходжкином (A. L. Hodg-kin) и А. Ф. Хаксли (A. F. Huxley) в 1952. Полный ионный ток слагается из трёх составляющих: калиевого, натриевого и тока утечки. Когда потенциал мембраны сдвигается на пороговую величину j* (~ 20мВ), мембрана становится проницаемой для ионов Na + . Ионы Na + устремляются внутрь волокна, сдвигая мембранный потенциал, пока он не достигнет величины равновесного натриевого потенциала:
![]()
составляющего ~ 60 мВ. Поэтому полная амплитуда потенциала действия достигает ~ 120 мВ. К моменту достижения макс. потенциала в мембране начинает развиваться калиевая (и одновременно уменьшаться натриевая) . В результате натриевый ток сменяется на калиевый, направленный наружу. Этот ток соответствует уменьшению потенциала действия.
Установлены эмпирич. ур-ния для описания натриевого и калиевого токов. Поведение мембранного потенциала при пространственно однородном возбуждении волокна определяется ур-нием:
где С -
ёмкость мембраны, I
- ионный ток, слагающийся из калиевого, натриевого и тока утечки. Эти токи определяются пост. эдс j K , j Na и j l
и проводимостями g
K , g
Na и g l:
Величину g l
считают постоянной, проводимости g
Na и g
K описывают с помощью параметров m
, h
и п:
g
Na , g
K - постоянные; параметры т, h
и п
удовлетворяют линейным ур-ниям

Зависимость коэф. a.
и b от мембранного потенциала j (рис. 3) выбирают из условия наилучшего совпадения

Рис. 3. Зависимость коэффициентов
a
.
и
b
от мембран
ного потенциала.
расчётных и измеряемых кривых I
(t
). Этими же соображениями вызван выбор параметров. Зависимость стационарных значений т, h
и п
от мембранного потенциала приведена на рис. 4. Существуют модели с большим числом параметров. Т. о., мембрана нервного волокна представляет собой нелинейный ионный проводник, свойства к-рого существенно зависят от электрич. поля. Механизм генерации возбуждения изучен плохо. Ур-ния Ходжкина -Хаксли дают лишь удачное эмпирич. описание явления, за к-рым нет конкретной физ. модели. Поэтому важной задачей является изучение механизмов протекания электрич. тока через мембраны, в частности через управляемые элект- рич. полем ионные каналы.

Рис. 4. Зависимость стационарных значений т, h
и п
от мембранного потенциала.
Распространение Н. и.
Н. и. может распространяться вдоль волокна без затухания и с пост. скоростью. Это связано с тем, что необходимая для передачи сигнала энергия не поступает из единого центра, а черпается на месте, в каждой точке волокна. В соответствии с двумя типами волокон существуют два способа передачи Н. и.: непрерывный и сальтаторный (скачкообразный), когда импульс движется от одного перехвата Ранвье к другому, перепрыгивая через области миелиновой изоляции.
В случае немиелинизир. волокна мембранного потенциала j(x, t
)определяется ур-нием:
где С -
ёмкость мембраны, приходящаяся на единицу длины волокна, R -
сумма продольных (внутриклеточного и внеклеточного) сопротивлений на единицу длины волокна, I
- ионный ток, протекающий через мембрану волокна единичной длины. Электрич. ток I
является функционалом от потенциала j, к-рый зависит от времени t
и координаты х.
Эта зависимость определяется ур-ниями (2) -(4).
Вид функционала I
специфичен для биологически возбудимой среды. Однако ур-ние (5), если отвлечься от вида I
, имеет более общий характер и описывает многие физ. явления, напр. процесс горения. Поэтому передачу Н. и. уподобляют горению порохового шнура. Если в бегущем пламени процесс поджигания осуществляется за счёт теплопроводности, то в Н. и. возбуждение происходит при помощи т. н. локальных токов (рис. 5).

Рис. 5. Локальные токи, обеспечивающие распростра
нение нервного импульса.
Ур-ния Ходжкина - Хаксли для распространения Н. и. решались численно. Полученные решения вместе с накопленными эксперим. данными показали, что распространение Н. и. не зависит от деталей процесса возбуждения. Качеств. картину распространения Н. и. можно получить при помощи простых моделей, отражающих лишь общие свойства возбуждения. Такой подход позволил рассчитывать и форму Н. и. в однородном волокне, их изменение при наличии неоднород-ностей и даже сложные режимы распространения возбуждения в активных средах, напр. в сердечной мышце. Существует неск. матем. моделей подобного рода. Простейшая из них такова. Ионный ток, протекающий через мембрану при прохождении Н. и., является знакопеременным: вначале он течёт внутрь волокна, а потом наружу. Поэтому его можно аппроксимировать кусочно-постоянной ф-цией (рис. 2, г
). Возбуждение происходит, когда мембранный потенциал сдвигается на пороговую величину j*. В этот момент возникает ток, направленный внутрь волокна и равный по модулю j".
Спустя t" ток меняется на противоположный, равный j
". Эта продолжается в течение времени ~ t ". Автомодельное решение ур-ния (5) можно найти как ф-цию переменной t = х/
u,
где u -
скорость распространения Н. и. (рис. 2, б).
В реальных волокнах время t" достаточно велико, поэтому только оно определяет скорость u,
для к-рой справедлива ф-ла: ![]() . Учитывая, что j
" ~ ~d, R ~ d
2 и С
~ d,
где d -
диаметр волокна, находим в согласии с экспериментом, что u ~ d
1/2 .
Спомощью кусочно-постоянной аппроксимации находят форму потенциала действия.
. Учитывая, что j
" ~ ~d, R ~ d
2 и С
~ d,
где d -
диаметр волокна, находим в согласии с экспериментом, что u ~ d
1/2 .
Спомощью кусочно-постоянной аппроксимации находят форму потенциала действия.
Ур-ние (5) для распространяющегося Н. и. в действительности допускает два решения. Второе решение оказывается неустойчивым; оно даёт Н. и. со значительно меньшей скоростью и амплитудой потенциала. Наличие второго, неустойчивого, решения имеет аналогию в теории горения. При распространении пламени с боковым теплоотводом также возможно возникновение неустойчивого режима. Простую аналитич. модель Н. и. можно усовершенствовать, учитывая дополнит. детали.
При изменении сечения и при ветвлении нервных волокон прохождение Н. и. может быть затруднено или даже полностью блокировано. В расширяющемся волокне (рис. 6) скорость импульса по мере приближения к расширению убывает, а после расширения начинает расти, пока не выйдет на новое стационарное значение. Замедление Н. и. тем сильнее, чем больше разница в сечениях. При достаточно большом расширении Н. и. останавливается. Существует критич. расширение волокна, к-рое задерживает Н. и.
При обратном движении Н. и. (из широкого волокна в узкое) блокирования не происходит, но изменение скорости носит противоположный характер. При подходе к сужению скорость Н. и. увеличивается, а затем начинает спадать до нового стационарного значения. На графике скорости (рис., 6 а
) получается своего рода петля гистерезиса.
Рие. 6. Прохождение нервных импульсов по расширя
ющемуся волокну: а -
изменение скорости импульса в
зависимости от его направления; б
-схематическое
изображение расширяющегося волокна.

Другой тип неоднородности - ветвление волокон. В узле ветвления возможны разл. варианты прохождения и блокирования импульсов. При несинхронном подходе Н. и. условие блокирования зависит от временного сдвига. Если временной между импульсами мал, то они помогают друг другу проникнуть в широкое третье волокно. Если сдвиг достаточно велик, то Н. и. мешают друг другу. Связано это с тем, что Н. и., подошедший первым, но не сумевший возбудить третье волокно, частично переводит узел в рефракторное состояние. Кроме того, возникает эффект синхронизации: по мере приближения Н. и. к узлу их запаздывание друг относительно друга уменьшается.
Взаимодействие Н. и.
Нервные волокна в организме объединены в пучки или нервные стволы, образующие подобие многожильного кабеля. Все волокна в пучке представляют собой самостоят. линии связи, но имеют один общий "провод" - межклеточную . Когда по любому из волокон бежит Н. и., он создаёт в межклеточной жидкости электрич. , к-рое влияет на мембранный потенциал соседних волокон. Обычно такое влияние пренебрежимо мало и линии связи работают без взаимных помех, но оно проявляется в пато-логич. и искусств. условиях. Обрабатывая нервные стволы спец. хим. веществами, удаётся наблюдать не только взаимные помехи, но и передачу возбуждения в соседние волокна.
Известны эксперименты по взаимодействию двух нервных волокон, помещённых в ограниченный объём внеш. раствора. Если по одному из волокон бежит Н. и., то одновременно изменяется возбудимость второго волокна. Изменение проходит три стадии. Вначале возбудимость второго волокна падает (повышается порог возбуждения). Это уменьшение возбудимости опережает потенциал действия, бегущий по первому волокну, и длится примерно до тех пор, пока потенциал в первом волокне не достигнет максимума. Затем возбудимость растёт, эта стадия совпадает по времени с процессом уменьшения потенциала в первом волокне. Возбудимость ещё раз уменьшается, когда в первом волокне происходит небольшая гиперполяризация мембраны.
При одноврем. прохождении Н. и. по двум волокнам иногда удавалось достигнуть их синхронизации. Несмотря на то что собств. скорости Н. и. в разных волокнах различны, при их одноврем. возбуждении мог возникнуть коллективный Н. и. Если собств. скорости были одинаковы, то коллективный импульс имел меньшую скорость. При заметном отличии собств. скоростей коллективная скорость имела промежуточное значение. Синхронизоваться могли лишь Н. и., скорости к-рых отличались не слишком сильно.
Матeм. описание этого явления даётся системой ур-ний для мембранных потенциалов двух параллельных волокон j 1 и j 2:

где R
1 и R
2 - продольные сопротивления первого и второго волокон, R
3 -
продольное сопротивление внешней среды, g = R
1 R
2 + R
1 R
3 .
+ R
2 R
3 .
Ионные токи I
1 и I
2 можно описать той или иной моделью нервного возбуждения.
При использовании простой аналитич. модели решение приводит к след. картине. Когда возбуждается одно волокно, в соседнем наводится знакопеременный мембранный потенциал: вначале волокно гиперполяри-зуется, затем деполяризуется и, наконец, ещё раз ги-перполяризуется. Эти три фазы соответствуют понижению, повышению и новому понижению возбудимости во-локна. При нормальных значениях параметров сдвиг мембранного потенциала во второй фазе в сторону деполяризации не достигает порога, поэтому передачи возбуждения в соседнее волокно не происходит. При одноврем. возбуждении двух волокон система (6) допускает совместное автомодельное решение, к-рое соответствует двум Н. и., движущимся с одинаковой скоростью на пост. расстоянии друг от друга. Если впереди находится медленный Н. и., то он притормаживает быстрый импульс, не выпуская его вперёд; оба движутся с относительно малой скоростью. Если же впереди находится быстрый II. и., то он подтягивает за собой медленный импульс. Коллективная скорость оказывается близкой к собств. скорости быстрого импульса. В сложных нейронных структурах возможно появление автоволи.
Возбудимые среды.
Нервные клетки в организме объединены в нейронные сети, к-рые в зависимости от частоты ветвления волокон разделяют на редкие и густые. В редкой сети отд. возбуждаются независимо друг от друга и взаимодействуют только в узлах ветвления, как описано выше.
В густой сети возбуждение охватывает сразу много элементов, так что их детальная структура и способ соединения между собой оказываются несущественными. Сеть ведёт себя как непрерывная возбудимая среда, параметры к-рой определяют возникновение и распространение возбуждения.
Возбудимая среда может быть трёхмерной, хотя чаще её рассматривают как двумерную . Возбуждение, возникшее в к.-л. точке поверхности, распространяется во все стороны в виде кольцевой волны. Волна возбуждения может огибать препятствия, но не может от них отражаться, не отражается она и от границы среды. При столкновении волн между собой происходит их взаимное уничтожение; пройти друг сквозь друга эти волны не могут из-за наличия позади фронта возбуждения рефракторной области.
Примером возбудимой среды является сердечный нервно-мышечный синцитий - объединение нервных и мышечных волокон в единую проводящую систему, способную передавать возбуждение в любом направлении. Нервно-мышечные синцитии сокращаются синхронно, подчиняясь волне возбуждения, к-рую посылает единый управляющий центр - водитель ритма. Единый ритм иногда нарушается, возникают аритмии. Один из таких режимов наз. трепетанием предсердий: это автономные сокращения, вызванные циркуляцией возбуждения вокруг препятствия, напр. верхней или нижней вены. Для возникновения подобного режима периметр препятствия должен превышать длину волны возбуждения, равную в предсердии человека ~ 5 см. При трепетании происходит пе-риодич. сокращение предсердий с частотой 3-5 Гц. Более сложный режим возбуждения представляет собой фибрилляция желудочков сердца, когда отд. элементы сердечной мышцы начинают сокращаться без внеш. команды и без связи с соседними элементами с частотой ~ 10 Гц. Фибрилляция приводит к прекращению циркуляции крови.
Возникновение и поддержание спонтанной активности возбудимой среды неразрывно связаны с возникновением источников волн. Простейший источник волн ( спонтанно возбуждающихся клеток) может обеспечить периодич. пульсацию активности, так устроен водитель ритма сердца.
Источники возбуждения могут возникать и за счёт сложной пространств. организации режима возбуждения, напр. ревербератор типа вращающейся спиральной волны, появляющийся в простейшей возбудимой среде. Другой вид ревербератора возникает в среде, состоящей из элементов двух типов с разными порогами возбуждения; ревербератор периодически возбуждает то одни, то другие элементы, меняя при этом направление своего движения и порождая плоские волны.
Третий вид источника - ведущий центр (источник эха), к-рый появляется в среде, неоднородной по реф-ракторности или порогу возбуждения. В этом случае на неоднородности возникает отражённая волна (эхо). Наличие подобных источников волн приводит к появлению сложных режимов возбуждения, исследуемых в теории автоволн.
Лит.: Ходжкин А., Нервный импульс, пер. с англ., М., 1965; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969; Тасаки И., Нервное возбуждение, пер. с англ., М., 1971; Маркин В. С., Пастушенко В. Ф., Чизмад-жев Ю. А., Теория возбудимых сред, М., 1981. В. С. Маркин.
НEРНСТА ТЕОРЕМА
- то же, что Третье начало термодинамики.
НEРНСТА ЭФФЕКТ (продольный гальванотермомаг-нитный эффект) - появление в проводнике, по к-рому течёт ток j , находящемся в магн. поле H | j , градиента темп-ры Т , направленного вдоль тока j ; градиент темп-ры не меняет знак при изменении направления поля Н на обратное (чётный эффект). Открыт В. Г. Нерн-стом (W. Н. Nernst) в 1886. Н. э. возникает в результате того, что перенос тока (поток носителей заряда) сопровождается потоком тепла. Фактически Н. э. представляет собой Пельтъе эффект в условиях, когда возникающая на концах образца разность темп-р приводит к компенсации потока тепла, связанного с током j , потоком тепла за счёт теплопроводности. Н. э. наблюдается также и в отсутствие магн. поля.
НEРНСТА-ЭТТИНГСХАУЗЕНА ЭФФЕКТ
- появление электрич. поля E
нэ в проводнике, в к-ром есть градиент темп-ры Т
,
в направлении, перпендикулярном магн. полю Н
.
Различают поперечный и продольный эффекты.
Поперечный H.-Э. э.
состоит в появлении электрич. поля Е
нэ |
(разности потенциалов V
нэ |
) в направлении, перпендикулярном Н
и Т
. В отсутствие магн. поля термоэлектрич. поле компенсирует поток носителей заряда, создаваемый градиентом темп-ры, причём компенсация имеет место лишь для полного тока: электроны с энергией, большей средней (горячие), движутся от горячего конца образца к холодному, электроны с энергией, меньшей средней (холодные),- в противоположном направлении. Сила Лоренца, отклоняет эти группы носителей в направлении, перпендикулярном Т
и магн. полю, в разные стороны; угол отклонения (угол Холла) определяется временем релаксации т данной группы носителей, т. е. различается для горячих и холодных носителей, если t зависит от энергии. При этом токи холодных и горячих носителей в поперечном направлении ( |
Т
и |
Н
) не могут компенсировать друг друга. Это приводит к появлению поля Е |
нэ ,
величина к-рого определяется из условия равенства 0 суммарного тока j
= 0.
Величина поля Е |
нэ зависит от Т, Н
и свойств вещества, характеризующихся коэф. Нернста-Эттингсха-узена N
|
:
![]()
В полупроводниках
под действием Т
носители заряда разных знаков движутся в одну сторону, а в магн. поле отклоняются в противоположные стороны. В результате направление поля Нернста - Эттингсхаузена, создаваемого зарядами разного знака, не зависит от знака носителей. Это существенно отличает поперечный Н.-Э. э. от Холла эффекта,
где направление поля Холла различно для зарядов разного знака.
Т. к. коэф. N |
определяется зависимостью времени т релаксации носителей от их энергии , то Н.-Э. э. чувствителен к механизму рассеяния носителей заряда.
Рассеяние носителей заряда уменьшает влияние магн. поля. Если t ~ , то при r
> 0 горячие носители рассеиваются реже холодных и направление поля Е |
нэ определяется направлением отклонения в магн. поле горячих носителей. При r
< 0 направление Е |
нэ противоположно и определяется холодными носителями.
В металлах,
где ток переносится электронами с энергией в интервале ~ kT
вблизи Ферми поверхности,
величина N |
задаётся производной д
t/д
.
на Ферми-поверхности = const (обычно у металлов N |
> 0, но, напр., у меди N |
< 0).
Измерения Н.-Э. э. в полупроводниках позволяют определить r,
т. е. восстановить ф-цию t(). Обычно при высоких темп-pax в области собств. проводимости полупроводника N |
<
0 из-за рассеяния носителей на оп-тич. фононах. При понижении темп-ры возникает область с N |
> 0, соответствующая примесной проводимости и рассеянием носителей гл. обр. на фононах (r
< < 0). При ещё более низких Т
доминирует рассеяние на ионизов. примесях с N |
< 0 (r
> 0).
В слабых магн. полях (w с t << 1, где w с - циклотронная частота
носителей) N |
не зависит от H
. В сильных полях (w c
t >> 1) коэф. N |
пропорц. 1/H
2 . В анизотропных проводниках коэф. N |
-
тензор. На величину N |
влияют увлечение электронов фотонами (увеличивает N |
),
анизотропия Ферми-поверхности и др.
Продольный H.
- Э. э.
состоит в возникновении элект-рич. поля Е ||
нэ (разности потенциалов V ||
нэ) вдоль Т
при наличии H
|
Т
. Т. к. вдоль Т
существует тер-моэлектрич. поле Е
a =
a Т
,
где a - коэф. термоэлек-трич. поля, то возникновение дополнит. поля вдоль Т
равносильно изменению поля Е
a .
при наложении магн. поля:

Магн. поле, искривляя траектории электронов (см. выше), уменьшает их длину свободного пробега l в направлении T . Т. к. время свободного пробега (время релаксации t) зависит от энергии электронов , то уменьшение l неодинаково для горячих и холодных носителей: оно меньше для той группы, для к-рой т меньше. Т. о., магн. поле меняет роль быстрых и медленных носителей в переносе энергии, и термоэлектрич. поле, обеспечивающее отсутствие заряда при переносе энергии, должно измениться. При этом коэф. N || также зависит от механизма рассеяния носителей. Термоэлектрич. ток растёт, если т падает с ростом энергии носителей (при рассеянии носителей на аку-стич. фононах), или уменьшается, если т увеличивается с увеличением (при рассеянии на примесях). Если электроны с разными энергиями имеют одинаковое t, эффект исчезает (N || = 0). Поэтому в металлах, где диапазон энергий электронов, участвующих в процессах переноса, мал (~ kT), N || мало: В полупроводнике с двумя сортами носителей N || ~ ~ g/kT. При низких темп-pax N || может также возрастать из-за влияния увлечения электронов фононами. В сильных магн. полях полное термоэлектрич. поле в магн. поле "насыщается" и не зависит от механизма рассеяния носителей. В ферромагн. металлах Н.-Э. э. имеет особенности, связанные с наличием спонтанной намагниченности.
Волна возбуждения, распространяющаяся по нервному волокну и проявляющаяся в электрич. (потенциал действия), ионных, механич., термич. и др. изменениях. Обеспечивает передачу информации от периферич. рецепторных окончаний к нервным центрам внутри… … Биологический энциклопедический словарь
Нервный импульс - См. Потенциал действия. Психология. А Я. Словарь справочник / Пер. с англ. К. С. Ткаченко. М.: ФАИР ПРЕСС. Майк Кордуэлл. 2000 … Большая психологическая энциклопедия
Нервный импульс электрический импульс, распространяющийся по нервному волокну. При помощи передачи нервных импульсов происходит обмен информацией между нейронами и передача информации от нейронов к клеткам других тканей организма. Нервный… … Википедия
Волна возбуждения, распространяющаяся по нервному волокну, в ответ на раздражение нейронов. Обеспечивает передачу информации от рецепторов в центральную нервную систему и от неё к исполнительным органам (мышцам, железам). Проведение нервного… … Энциклопедический словарь
Нервный импульс - волна возбуждения, которая распространяется вдоль нервных волокон и по телу нервных клеток в ответ на раздражение нейронов и служит для передачи сигнала от рецепторов в центральную нервную систему, а от нее к исполнительным органам (мышцам,… … Начала современного естествознания
нервный импульс - nervinis impulsas statusas T sritis Kūno kultūra ir sportas apibrėžtis Jaudinimo banga, plintanti nerviniu audiniu. Atsiranda padirginus nervų ląsteles. Perduoda signalus iš jautriųjų periferinių nervų galūnių (receptorių) į centrinę nervų… … Sporto terminų žodynas
См. Импульс нервный … Большая советская энциклопедия
НЕРВНЫЙ ИМПУЛЬС - См. импульс (4) … Толковый словарь по психологии
Проведение нервных импульсов по нервным волокнам и через синапсы. Высоковольтный потенциал, возникающий при возбуждении рецептора в нервном волокне, в 5-10 раз больше порога раздражения рецептора. Проведение волны возбуждения по нервному волокну обеспечивается тем, что каждый последующий его участок раздражается высоковольтным потенциалом предыдущего участка. В мякотных нервных волокнах этот потенциал распространяется не непрерывно, а скачкообразно; он перескакивает через один или даже несколько перехватов Ранвье, в которых усиливается. Продолжительность проведения возбуждения между двумя соседними перехватами Ранвье равняется 5-10% длительности высоковольтного потенциала.
Проведение нервного импульса по нервному волокну происходит только при условии его анатомической непрерывности и нормального физиологического его состояния. Нарушение физиологических свойств нервного волокна сильным охлаждением или отравлением ядами и наркотиками прекращает проведение нервного импульса даже при анатомической его непрерывности.
Нервные импульсы проводятся изолированно по отдельным двигательным и чувствительным нервным волокнам, которые входят в состав смешанного нерва, что зависит от изолирующих свойств покрывающих их миелиновых оболочек. В безмякотных нервных волокнах биоток распространяется непрерывно вдоль волокна и благодаря соединительнотканой оболочке не переходит с одного волокна на другое. Нервный импульс может распространяться по нервному волокну в двух направлениях: центростремительном и центробежном. Следовательно, существуют три правила проведения нервного импульса в нервных волокнах: 1) анатомической непрерывности и физиологической целости, 2) изолированного проведения и 3) двустороннего проведения.
Через 2-3 дня после отделения нервных волокон от тела нейрона они начинают перерождаться, или дегенерировать, и проведение нервных импульсов прекращается. Нервные волокна и миелин разрушаются и сохраняется только соединительнотканая оболочка. Если соединить перерезанные концы нервных волокон, или нерва, то после дегенерации тех участков, которые отделены от нервных клеток, начинается восстановление, или регенерация, нервных волокон со стороны тел нейронов, из которых они прорастают в сохранившиеся соединительнотканые оболочки. Регенерация нервных волокон приводит к восстановлению проведения импульсов.
В отличие от нервных волокон через нейроны нервной системы нервные импульсы проводятся только в одном направлении - от рецептора к работающему органу. Это зависит от характера проведения нервного импульса через синапсы. В нервном волокне над пресинаптической мембраной есть множество мельчайших пузырьков ацетилхолина. При достижении биотоком пресинаптической мембраны часть этих пузырьков лопается, и ацетилхолин проходит через мельчайшие отверстия в пресинаптической мембране в синаптическую щель.
В постсинаптической мембране имеются участки, обладающие особым сродством к ацетилхолину, который вызывает временное появление пор в постсинаптической мембране, отчего она становится временно проницаемой для ионов. В результате в постсинаптической мембране возникает возбуждение и высоковольтный потенциал, который распространяется по следующему нейрону или по иннервируемому органу. Следовательно, передача возбуждения через синапсы происходит химическим путем посредством посредника, или медиатора, ацетилхолина, а проведение возбуждения по следующему нейрону снова осуществляется электрическим путем.
Действие ацетилхолина на проведение нервного импульса через синапс кратковременно; он быстро разрушается, гидролизуется ферментом холинэстеразой.
Так как химическая передача нервного импульса в синапсе происходит в течение доли мсек, то в каждом синапсе нервный импульс на это время задерживается.
В отличие от нервных волокон, в которых информация передается по принципу «все или ничего», т. е. дискретно, в синапсах информация передается по принципу «больше или меньше», т. е. градуально. Чем больше до некоторого предела образуется медиатора ацетилхолина, тем выше частота высоковольтных потенциалов в последующем нейроне. После этого предела возбуждение переходит в торможение. Таким образом, цифровая информация, передаваемая по нервным волокнам, переходит в синапсах в измерительную информацию. Измерительные электронные машины,
в которых имеются определенные соотношения между реально измеряемыми количествами и теми величинами, которые они представляют, называются аналоговыми, работающими по принципу «больше или меньше»; можно считать, что в синапсах происходит аналогичный процесс и совершается его переход в цифровой. Следовательно, нервная система функционирует по смешанному типу: в ней совершаются и цифровые и аналоговые процессы.