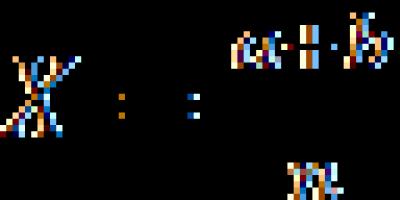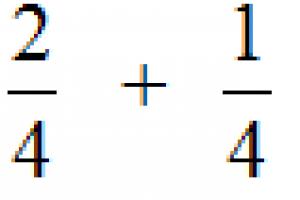Хромосомный уровень организации наследственного материала. Хромосомы, как группы сцепления генов.
Из принципов генетического анализа вытекает, что независимое комбинирование признаков может осуществляться лишь при условии, что гены, определяющие эти признаки, находятся в разных парах хромосом. Следовательно, у каждого организма, число пар признаков, по которым наблюдается независимое наследование, ограничено числом пар хромосом. С другой стороны, очевидно, что число признаков и свойств организма, контролируемых генами, чрезвычайно велико, а число пар хромосом у каждого вида относительно мало и постоянно. Остается допустить, что в каждой хромосоме находится не один ген, а много. Если это так, то следует признать, что третье правило Менделя касается только распределения хромосом, а не генов, т.е. его действие ограничено. Анализ проявления третьего правила показал, что в некоторых случаях новые комбинации генов у гибридов совсем отсутствовали, т.е. наблюдалось полное сцепление между генами исходных форм и в фенотипе наблюдалось расщепление 1:1. В других случаях комбинация признаков отмечалась с меньшей частотой, чем ожидается при независимом наследовании.
В 1906 году У. Бетсон описал нарушение менделевского закона независимого наследования двух признаков. Возникли вопросы: почему не все признаки наследуются и как они наследуются, как расположены гены в хромосомах, каковы закономерности наследования генов, находящихся в одной хромосоме? На эти вопросы смогла ответить хромосомная теория наследственности, созданная Т. Морганом, в 1911 году.
Т. Морган, изучив все отклонения, предложил называть совместное наследование генов, ограничивающее их свободное комбинирование, сцеплением генов или сцепленным наследованием.
Закономерности полного и неполного сцепления. Группы сцепления у человека.
Исследования Т. Моргана и его школы показали, что в гомологичной паре хромосом регулярно происходит обмен генами. Процесс обмена идентичными участками гомологичных хромосом с содержащимися в них генами называют перекрестом хромосом или кроссинговером. Кроссинговер наблюдается в мейозе. Он обеспечивает новые сочетания генов, находящихся в гомологичных хромосомах. Явление кроссинговера, как и сцепление генов, характерно для животных, растений, микроорганизмов. Исключение составляют самцы дрозофилы и самки тутового шелкопряда. Кроссинговер обеспечивает рекомбинацию генов и тем самым значительно увеличивает роль комбинативной изменчивости в эволюции. О наличии кроссинговера можно судить на основе учета частоты возникновения организмов с новым сочетанием признаков. Явление кроссинговера было открыто Морганом на дрозофиле.
Запись генотипа дигетерозиготы при независимом наследовании:
А В
Запись генотипа дигетерозиготы при сцепленном наследовании:
Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными, а не претерпевшие – некроссоверными.
АВ, ав Ав, аВ
Некроссоверные гаметы. Кроссоверные гаметы.
Соответственно организмы, возникшие от сочетания кроссоверных гамет, называют кроссоверами или рекомбинантами, а возникшие от сочетания некроссоверных гамет – некроссоверами или нерекомбинантами .
Явление кроссинговера, как и сцепление генов, можно рассмотреть и в классическом опыте Т. Моргана при скрещивании дрозофил.
|
Признак |
P♀ BV x ♂ bv |
|
|
серый цвет тела |
||
|
черный цвет тела |
||
|
нормальные крылья |
||
|
рудиментарные крылья |
||
|
Анализирующее скрещивание 1. Полное сцепление генов. 2. Неполное сцепление генов. |
1. Полное сцепление P♀ bv x ♂ BV F 2 bv bv расщепление – 1:1 |
|
|
2. Неполное сцепление (кроссинговер) P:♀ BV x ♂ bv G: BV bv Bv bV bv некроссоверные кроссоверные F 2 BV bv Bv bV некроссоверов – 83% кроссоверов – 17% |
||
Для измерения расстояния между генами путем анализирующего скрещивания можно применять формулу:
![]() где:
где:
X – расстояние между генами в % кроссинговера или в морганидах;
а – количество особей 1-й кроссоверной группы;
в – количество особей 2-й кроссоверной группы;
n – общее количество гибридов в опыте;
100% – коэффициент для перевода в проценты.
На основании исследования сцепленного наследования Морган сформулировал тезис, вошедший в генетику под названием правило Моргана : гены, локализованные в одной хромосоме, наследуются сцеплено, причем сила сцепления зависит от расстояния между ними.
Сцепленные гены расположены в линейном порядке и частота кроссинговера между ними прямо пропорциональна расстоянию между ними. Однако, этот тезис характерен только для близко лежащих друг к другу генов. В случае же относительно удаленных генов наблюдается некоторое отклонение от такой зависимости.
Морган предложил выражать расстояние между генами в процентах кроссинговера между ними. Расстояние между генами также выражают в морганидах или сантиморганидах. Морганида – генетическое расстояние между генами, где происходит кроссинговер с частотой 1%.
По частоте кроссинговера между двумя генами можно судить об относительном расстоянии между ними. Так, если между генами А и В кроссинговер составляет 3%, а между генами В и С – 8% кроссинговера, то между А и С кроссинговер должен происходить с частотой либо 3+8=11%, либо 8-3=5%, в зависимости от того, в каком порядке эти гены расположены в хромосоме.
А ─ ─ ─ В ─ ─ ─ ─ ─ ─ ─ ─ С В ─ ─ ─ А ─ ─ ─ ─ ─ ─ ─ ─ С
Задача 1. Катаракта и полидактилия наследуются как доминантные аутосомные признаки. Женщина унаследовала катаракту от отца, полидактилию от матери. Гены сцеплены, расстояние между ними 3М. Каковы генотипы и фенотипы детей от брака этой женщины и мужчины нормального по этим признакам? Какова вероятность рождения здоровых детей?
|
катаракта |
P ♀ аВ х ♂ ав |
|
|
полидактилия |
Х = АВ = 3 Морг. |
|
|
P ♀ аВ х ♂ ав |
Ответ: вероятность рождения здорового ребенка – 1,5%, имеющих по 1 признаку – 48,5%, имеющих оба признака – 1,5% |
|
|
G: (аВ) (Ав) (ав) |
||
|
F1 аВ Ав ав АВ ав ав ав ав 48,5% 48,5% 1,5% 1,5% |
||
Генетическая карта хромосомы – это схема, отображающая порядок расположения генов на относительном расстоянии их друг от друга. О расстоянии между сцепленными генами судят по частоте кроссинговера между ними. Генетические карты всех хромосом составлены для наиболее изученных в генетическом отношении организмов: дрозофилы, кур, мышей, кукурузы, томатов, нейроспоры. Для человека также составлены генетические карты всех 23 хромосом.
После установления линейной дискретности хромосом возникла необходимость составления цитологических карт с целью сопоставления с генетическими, составленными на основе учета рекомбинаций.
Цитологическая карта – это карта хромосомы, на которой определяется расположение и относительное расстояние между генами в самой хромосоме. Построение их ведется на основе анализа хромосомных перестроек, дифференциальной окраски политенных хромосом, радиоактивных меток и др.
К настоящему времени, у ряда растений и животных построены и сопоставлены генетические и цитологические карты. Реальность этого сопоставления подтверждает правильность принципа о линейном расположении генов в хромосоме.
У человека можно назвать некоторые случаи сцепленного наследования.
Гены, контролирующие наследование групп крови по системе АВ0 и синдрома дефекта ногтей и коленной чашечки, наследуются сцепленно.
Сцеплены гены резус-фактора и овальной формы эритроцитов.
В третьей аутосоме расположены гены группы крови Лютеран и секреции антигенов А и В со слюной.
Гены полидактилии и катаракты наследуются сцепленно.
В Х-хромосоме расположены гены гемофилии и дальтонизма, а также гены цветовой слепоты и мышечной дистрофии Дюшена.
В 6 аутосоме находятся сублокусы А, В, С, D/DR системы HLA, контролирующих синтез антигенов гистосовместимости.
Наследование признаков Х-сцепленных и голандрических.
Признаки, контролируемые генами, расположенными в половых хромосомах, называются сцепленным с полом. У человека описано более 60 заболеваний, сцепленных с полом, большинство из которых наследуются рецессивно. Гены в половых хромосомах можно разделить на 3 группы:
Гены частично сцепленные с полом. Они расположены в парных сегментах Х и Y хромосом . К заболеваниям частично сцепленным с полом относят: геморрагический диатез, судорожные расстройства, пигментный ретинит, пигментную ксеродерму, общую цветовую слепоту.
Гены полностью сцепленные с полом. Они расположены в участке Х хромосомы , для которого нет гомологичного участка в Y хромосоме (гетерологическом). Эти гены контролируют заболевания: атрофия зрительного нерва, мышечная дистрофия Дюшена, дальтонизм, гемофилия, способность ощущать запах синильной кислоты.
Гены, расположенные в участке Y хромосомы , для которого нет гомологичного локуса в Х хромосоме, называются голандрическими . Они контролируют признаки: синдактилия, гипертрихоз ушной раковины.
Ген дальтонизма проявляется у 7% мужчин и у 0,5% женщин, но носительницами этого гена являются 13% женщин.
Сцепленное с полом наследование было описано Т. Морганом на примере наследования признака окраски глаз у дрозофилы.
Отмечено несколько закономерностей наследования сцепленных с полом признаков:
передаются крест на крест (от отца – дочери, от матери – сыну);
результаты прямого и обратного скрещиваний не совпадают;
у гетерогаметного пола признак проявляется в любом состоянии (доминантном или рецессивном).
Основные положения хромосомной теории наследственности.
Основные положения хромосомной теории наследственности можно сформулировать следующим образом:
Гены находятся в хромосомах. Каждый ген в хромосоме занимает определенный локус. Гены в хромосомах расположены линейно.
Каждая хромосома представляет группу сцепленных генов. Число групп сцепления у каждого вида равно числу пар хромосом.
Между гомологичными хромосомами происходит обмен аллельными генами – кроссинговер.
Расстояние между генами в хромосоме пропорционально проценту кроссинговера между ними. Зная расстояние между генами можно вычислить процентное соотношение генотипов у потомства.
Сцепленное наследование - наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Вместе с признаками, которые наследуются независимо, должны существовать и такие, которые наследуются сцеплено друг с другом, так как они определяются генами, расположенными в одной хромосоме. Такие гены образуют группу сцепления . Количество групп сцепления в организмах определенного вида равно количеству хромосом в гаплоидном наборе (например, у дрозофилы 1пара = 4, у человека 1пара = 23).
Полное сцепление - разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Неполное сцепление - разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
(Кроссоверные гаметы - гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
Кроссинговер - обмен участками гомологичных хромосом в процессе клеточного деления, преимущественно в профазе первого мейотического разделения, иногда в митозе. Опытами Т. Моргана, К. Бриджеса и А. Стертеванта было показано, что нет абсолютно полного сцепления генов, при котором гены передавались бы всегда вместе. Вероятность того, что два гена, локализованные в одной хромосоме, не разойдутся в процессе мейоза, колеблется в пределах 1-0,5. В природе преобладает неполное сцепление, обусловленное перекрёстком гомологичных хромосом и рекомбинацией генов. Цитологическая картина кроссинговера была впервые описана датским ученым Ф. Янсенсом.
Кроссинговер проявляется только тогда, когда гены находятся в гетерозиготном состоянии (АВ / ав). Если гены находятся в гомозиготном состоянии (АВ / АВ или аВ/аВ), обмен идентичными участками не дает новых комбинаций генов в гаметах и в поколении. Частота (процент) перекрёстка между генами зависит от расстояния между ними: чем дальше они располагаются друг от друга, тем чаще происходит кроссинговер. Т. Морган предложил расстояние между генами измерять кроссинговером в процентах, по формуле:
N1/N2 X 100 = % кроссинговера ,
где N1 - общее число особей в F;
N2 - суммарное число кроссоверных особей.
Отрезок хромосомы, на котором осуществляется 1% кроссинговера, равна одной морганиде (условная мера расстояния между генами). Частоту кроссинговера используют для того, чтобы определить взаимное расположение генов и расстояние между ними. Для построения генетической карты человека пользуются новыми технологиями, кроме того построены цитогенетические карты хромосом.
Различают несколько типов кроссинговера: двойной, множественный (сложный), неправильный, неровный.
Кроссинговер приводит к новому сочетанию генов, вызывает изменение фенотипа. Кроме того, он наряду с мутациями является важным фактором эволюции организмов.)
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
· гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
· каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
· гены расположены в хромосомах в определенной линейной последовательности;
· гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
· сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
· каждый вид имеет характерный только для него набор хромосом - кариотип.
Наследование пола и признаков, сцепленных с полом. Половые хромосомы и их роль в детерминации пола.Наследование пола. Пол особи - это сложный признак, формируемый как действием генов, так и условиями развития. У человека одна из 23 пар хромосом - половые хромосомы, обозначаемые как X и Y. Женщины - гомогаметный пол, т.е. имеют две X-хромосомы, одну - полученную от матери, а другую - от отца. Мужчины - гетерогаметный пол, имеют одну X- одну Y-хромосому, причем X передается от матери, а Y - от отца. Заметим, что гетерогаметный пол не всегда обязательно мужской; например, у птиц это самки, в то время как самцы гомогаметны. Имеются и другие механизмы детерминации пола. Так, у ряда насекомых Y-хромосома отсутствует. При этом один из полов развивается при наличии двух X-хромосом, а другой - при наличии одной X-хромосомы. У некоторых насекомых пол определяется соотношением числа аутосом и половых хромосом. У ряда животных может происходить т.н. переопределение пола, когда в зависимости от факторов внешней среды зигота развивается либо в самку, либо в самца. Развитие пола у растений имеет столь же разнообразные генетические механизмы, как и у животных.
Признаки, сцепленные с X-хромосомой . Если ген находится в половой хромосоме (его называют сцепленным с полом), то проявление его у потомков следует иным, чем для аутосомых генов, правилам. Рассмотрим гены, находящиеся в X-хромосоме. Дочь наследует две X-хромосомы: одну - от матери, а другую - от отца. Сын же имеет только одну X-хромосому - от матери; от отца же он получает Y-хромосому. Поэтому отец передает гены, имеющиеся в его X-хромосоме, только своей дочери, сын же их получить не может. Поскольку X-хромосома более "богата" генами по сравнению с Y-хромосомой, то в этом смысле дочь генетически более схожа с отцом, чем сын; сын же более схож с матерью, чем с отцом.
Один из исторически наиболее известных сцепленных с полом признаков у человека - это гемофилия, приводящая к тяжелым кровотечениям при малейших порезах и обширным гематомам при ушибах. Она вызывается рецессивным дефектным аллелем 0, блокирующим синтез белка, необходимого для свертывания крови. Ген этого белка локализован в Х-хромосоме. Гетерозиготная женщина +0 (+ означает нормальный активный аллель, доминантный по отношению к аллелю гемофилии 0) не заболевает гемофилией, и ее дочери тоже, если у отца нет этой патологии. Однако ее сын может получить аллель 0, и тогда у него развивается гемофилия. Рецессивные заболевания, вызываемые генами X-хромосомы, намного реже поражают женщин, чем мужчин, поскольку у них заболевание проявляется только при гомозиготности - наличии рецессивного аллеля в каждой из двух гомологичных X-хромосом; мужчины заболевают во всех случаях, когда их единственная X-хромосома несет дефектный аллель.
Сцепление с Y-хромосомой. Сведения о генах, находящихся в Y-хромосоме, весьма скудны. Предполагается, что она практически не несет генов, обусловливающих синтез белков, необходимых для функционирования клетки. Но она играет ключевую роль в развитии мужского фенотипа. Отсутствие Y-хромосомы при наличии только одной X-хромосомы приводит к т.н. синдрому Тернера: развитию женского фенотипа с плохо развитыми первичными и вторичными половыми признаками и другими отклонениями от нормы. Встречаются мужчины с добавочной Y-хромосомой (XYY); они высокого роста, агрессивны и нередко аномального поведения. В Y-хромосоме выявлено несколько генов, ответственных за регуляцию синтеза специфических ферментов и гормонов, и нарушения в них приводят к патологиям полового развития. Имеется ряд морфологических признаков, которые, как полагают, определяются генами Y-хромосомы; среди них - развитие волосяного покрова ушей. Подобного рода признаки передаются только по мужской линии: от отца к сыну.
Генетическая детерминация пола, определяемая набором половых хромосом, поддерживает равное воспроизводство самок и самцов. Действительно, женские яйцеклетки содержат только X-хромосому, поскольку женщины имеют генотип XX по половым хромосомам. Генотип же мужчин - XY, и потому рождение девочки или мальчика в каждом конкретном случае определяется тем, несет ли спермий X- или Y-хромосому. Поскольку же в процессе мейоза хромосомы имеют равные шансы попасть в гамету, то половина гамет, производимых индивидами мужского пола, содержит X-, а половина - Y-хромосому. Поэтому половина потомков ожидается одного пола, а половина - другого.
Следует подчеркнуть, что предсказать заранее рождение мальчика или девочки невозможно, поскольку невозможно предугадать, какая мужская половая клетка будет участвовать в оплодотворении яйцеклетки: несущая X- или Y-хромосому. Поэтому наличие большего или меньшего числа мальчиков в семье - дело случая.
Биологической основой третьего закона Менделя является независимое расхождение хромосом при мейозе. Поэтому третий закон верен только для генов, находящихся в разных хромосомах.
Если гены находятся в одной хромосоме, то они не могут разойтись независимо друг от друга, поэтому наследуются вместе (сцеплено) - это закон сцепления (закон Моргана).Все гены, находящиеся в одной хромосоме, образуют группу сцепления.
При полном сцеплении (встречается, например, у самцов дрозофил) дигетерозигота образует только два типа гамет.
Гораздо чаще встречается неполное сцепление, когда из-за кроссинговера при мейозе происходит обмен участками хромосом. Тогда дигетерозигота образует 4 типа гамет в неравном соотношении: большую часть составляют гаметы с группой сцепления, меньшую - рекомбинантные гаметы.
Доля рекомбинантных гамет зависит от расстояния между генами в хромосоме, измеряется в условных единицах морганидах. Фраза «расстояние между генами А и В равняется 10 морганид» означает, что рекомбинантных гамет получится в сумме 10% (5%+5%), а нормальных - 90% (45% и 45%).
Тесты
1. При скрещивании мух дрозофил с серым телом и нормальными крыльями и дрозофил с темным телом и зачаточными крыльями проявляется закон сцепленного наследования, следовательно, эти гены расположены в
А) разных хромосомах и сцеплены
Б) одной хромосоме и сцеплены
В) одной хромосоме и не сцеплены
Г) разных хромосомах и не сцеплены
2. Если гены расположены в разных парах негомологичных хромосом, то проявляется закон
А) неполного доминирования
Б) полного доминирования
В) независимого наследования
Г) расщепления признаков
3. Если гены, ответственные за окраску и форму семян гороха, расположены в разных хромосомах, то во втором поколении проявляется закон
А) независимого наследования
Б) сцепленного наследования
В) расщепления признаков
Г) доминирования
4. Количество групп сцепления генов у организмов зависит от числа
А) пар гомологичных хромосом
Б) аллельных генов
В) доминантных генов
Г) молекул ДНК в ядре клетки
5. Если гены, отвечающие за развитие нескольких признаков, расположены в одной хромосоме, то проявляется закон
А) расщепления
Б) сцепленного наследования
В) неполного доминирования
Г) независимого наследования
6. "Гены, расположенные в одной хромосоме, наследуются совместно" - это формулировка закона
А) взаимодействия генов
Б) сцепленного наследования
В) независимого наследования
Г) гомологических рядов изменчивости
7. Какой закон проявляется при скрещивании дигетерозиготных организмов, у которых гены, например А и В, расположены в негомологичных хромосомах?
А) полного доминирования
Б) неполного доминирования
В) независимого наследования
Г) сцепленного наследования
8. Всегда наследуются вместе гены
А) рецессивные
Б) аллельные
В) доминантные
Г) тесно сцепленные
9. При скрещивании дрозофил с серым телом и нормальными крыльями и дрозофил с темным телом и зачаточными крыльями проявляется закон сцепленного наследования, так как отвечающие за эти признаки гены расположены в
А) ДНК митохондрий
Б) разных парах хромосом
В) одной паре хромосом
Г) половых хромосомах
10. Какой закон проявится при скрещивании, если гены расположены в одной хромосоме?
А) расщепления признаков
Б) сцепленного наследования
В) независимого наследования
Г) гомологических рядов
11. Согласно закону Т. Моргана гены наследуются преимущественно вместе, если они расположены в
А) аутосоме
Б) половых хромосомах
В) одной хромосоме
Г) разных гомологичных хромосомах
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган . Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F 1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.

1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb , а отцовский — один тип — аb . Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb . Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb . Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В , появляются гаметы Аb и аВ , и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза ), или в разных (транс-фаза ).

1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности :
- гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
- каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
- гены расположены в хромосомах в определенной линейной последовательности;
- гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
- сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
- каждый вид имеет характерный только для него набор хромосом — кариотип.
Перейти к лекции №17 «Основные понятия генетики. Законы Менделя»
Вопрос 1. Что такое сцепленное наследование?
Сцепленное наследование
- это совместное наследование генов, находящихся в одной хромосоме (т. е. в одной молекуле ДНК). Например, у душистого горошка гены, определяющие окраску цветков и форму пыльцы, расположены именно таким образом. Они наследуются сцепленно, поэтому при скрещивании у гибридов второго поколения образуются родительские фенотипы в соотношении 3:1, а расщепление 9:3:3:1, характерное для дигибридного скрещивания при независимом наследовании, не проявляется.
При сцепленном наследовании сила сцепления может быть разной. При полном сцеплении в потомстве гибрида появляются организмы только с родительскими сочетаниями признаков, а рекомбинанты отсутствуют. При неполном сцеплении всегда наблюдается в той или иной мере преобладание форм с родительскими признаками. Величина кроссинговера, отражающая силу сцепления между генами, измеряется отношением числа рекомбинантов к общему числу в потомстве от анализирующего скрещивания и выражается в процентах.
Гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними. За единицу расстояния между двумя генами условно принимают 1% перекреста между ними - эту величину называют морганидой.
Чем дальше друг от друга расположены два гена в хромосомах, тем больше вероятности, что между ними произойдет кроссинговер. Следовательно, по частоте кроссинговера между генами можно судить об относительном расстоянии, разделяющим гены в хромосоме, при этом гены в хромосоме расположены в линейном порядке.
Каждая хромосома в кариотипе человека несет в себе множество генов, которые могут наследоваться совместно.
Вопрос 2. Что представляют собой группы сцепления генов?
Явление совместного наследования генов было впервые описано Пеннетом, назвавшим это явление “притяжением генов”. Томас Хант Морган и его сотрудники подробно изучили явление сцепленного наследования генов и вывели законы сцепленного наследования (1910). Группа сцепления – это совокупность генов, локализованных в одной хромосоме. Число групп сцеплений для каждого вида равно гаплоидному набору хромосом, а точнее – равно количеству пар гомологичный хромосом. У человека половая пара хромосом негомологична, поэтому у женщин групп сцепления – 23, а у мужчин – 24 (22 группы сцепления - аутосомные и две по половым хромосомам Х и У). У гороха 7 групп сцепления (2n = 14), у дрозофилы - 4 группы сцепления (2n = 8).
Вопрос 3. Что является причиной нарушения сцепления генов?
Причиной нарушения сцепления генов является обмен участками гомологичных хромосом в профазе I мейотического деления. Напомним, что на этом этапе парные хромосомы конъюгируют, образуя так называемые биваленты. Формирование бивалентов может привести к перекресту хромосом, что создает возможность обмена гомологичными участками ДНК. Если это происходит, то группы сцепления меняют свое содержание (в них оказываются иные аллели тех же генов) и в потомстве могут появиться особи с фенотипом, отличающимся от родительских.
Вопрос 4. Каково биологическое значение обмена аллельными генами между гомологичными хромосомами?
Кроссинговер – обмен идентичными участками между гомологичными хромосомами, приводящий к рекомбинации наследственных задатков и формированию новых сочетаний генов в группах сцепления.
Перекрест хромосом приводит к перекомбинированию генетического мтериала и формированию новых сочетаний аллелей генов из группы сцепления. При этом увеличивается разнообразие потомков, т. е. повышается наследственная изменчивость, что имеет большое эволюционное значение. Действительно, если, например, у дрозофилы гены, определяющие окраску тела и длину крыльев, находятся на одной хромосоме, то, скрещивая чистые линии серых мух с нормальными крыльями и черных мух с укороченными крыльями, в отсутствие крос-синговера мы никогда не получим иные фенотипы. Существование же перекреста хромосом позволяет появиться (в нескольких процентах случаев) серым мухам с короткими крыльями и черным мухам с нормальными крыльями.
Вопрос 5. Подтверждена ли цитологически теория сцепленного наследования?
Теория сцепленного наследования Томаса Ханта Моргана (1866-1945) подтверждена цитологическими наблюдениями. Было показано, что хромосомы при делении целиком расходятся к разным полюсам клетки. Следовательно, гены, расположенные на одной хромосоме, при мейозе попадают в одну гамету, т.е. действительно наследуются сцепленно.